



2022年12月,西南大学梁国鲁教授团队完成了野生枇杷高质量基因组和种质资源重测序研究,相关研究成果“Genome assembly of wild loquat (Eriobotrya japonica) and resequencing provide new insights into the genomic evolution and fruit domestication in loquat”发表于园艺学国际顶刊期刊《Horticulture Research》上。太阳集团电子游戏有幸参与了该研究,完成了其中基因组、群体和转录组测序及分析等工作。

研究背景
枇杷是世界受欢迎的果实之一,它与苹果、梨、草莓等经济物种同属于蔷薇科。枇杷起源于中国,已种植了2000多年,现已广泛分布在全球30多个国家,其水果含丰富的营养物质,包括糖类、氨基酸、维生素、有机酸和矿物质等。此外,其果实在食品工业中也常用来制作果汁、葡萄酒、糖浆和果酱。枇杷在进化和驯化历史上一直存在争议。目前,野生枇杷为系统进化和育种提供了大量宝贵的遗传资源,野生和栽培种质间的遗传变异变异研究有助于更好地了解作物物种的驯化过程,同时也为功能基因鉴定提供有效的途径,后期可以更好地应用于遗传改良。然而,由于复杂的遗传背景和悠久的栽培历史,野生枇杷和栽培枇杷之间的基因组进化规律仍不清楚。
材料方法
Denovo:~159.8 Nanopore reads(~198.5×),IIIlumina reads,Hi-C,~8G RNA-seq data基因组注释
重测序:26份栽培枇杷和11份野生枇杷,Illumina测序,~22.49×
转录组:果实未成熟(GF)、果实转色(CT)和果实成熟(FR)3个时期转录组分析
代谢组:Jinhua No. 2 FR与GZ-23 FR,Huabai No. 1 FR与GZ-White FR, LC-MS/MS广靶
研究结果
1、野生枇杷GZ-23基因组的组装及注释
作者对野生枇杷(Eriobotrya japonica Lindl.)(2n = 34)的基因组进行了研究,与流式细胞仪预估的结果(∼760Mb)相一致,k-mer分析结果显示基因组大小为737.06 Mb。随后,利用152.6 Gb的ONT数据(reads N50 = 31.6kb)进行基因组组装,同时利用二代数据纠错和Hi-C数据挂载,成功组装了分布在17条染色体上的783.7 Mb的野生枇杷基因组序列(图1);其中,scaffold N50和contig N50分别为41.8Mb和3.9Mb。二代转录组的回比率为98.7%,CEGMA和BUSCO评估分别为98.03%和98.27%,表明GZ-23基因组组装的完整性较好(表1)。进而结合从头预测、同源比对预测和转录组辅助预测,共预测到45,791个基因,鉴定到59.75%的基因组重复序列,此外,还鉴定出5381个rRNAs、765个tRNAs和129个miRNAs。
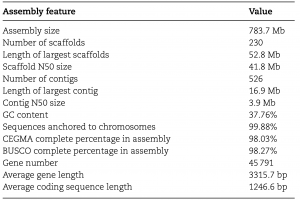
表1 野生枇杷基因组组装统计及注释
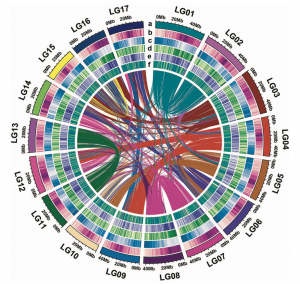
图1 野生枇杷基因组组装特征
2、野生枇杷与12个物种的比较基因组分析
为研究野生枇杷基因组特征及进化,作者将其与蓝星睡莲、水稻、铁皮石斛、番茄,茶树、猕猴桃、苹果、白梨、野草莓、拟南芥,杧果和葡萄基因组进行比较基因组分析,通过基因家族聚类,发现92%(42,150)的蛋白编码基因聚为20,668个基因家族,其中523个基因家族为枇杷特有的单拷贝(图2A),主要参生物学质量调控、激素水平、定位、运输和生长素极性运输等生物过程。通过4DTv和Ks对全基因组复制事件分析表明,枇杷基因组中发生了两次全基因组复制(WGD)事件,最近的WGD事件发生在30-45 Mya,而古WGD可能发生在已知的约120~130 Mya的古六倍化WGT(γ)事件。系统分化表明,枇杷与苹果和梨拥有共同的祖先,这三个物种的分化发生在最近的一次WGD事件之后,通过比较分析,枇杷基因组104个基因家族发生扩张,仅有一个基因家族发生收缩,GO富集分析表明,发生扩张的基因家族主要参与花粉识别、分生组织发育与维持、免疫反应、和DNA代谢过程的调节等生物过程;KEGG分析表明,主要参与半乳糖代谢、氨基酸代谢、氨基糖/核苷酸糖代谢、氮代谢、不饱和脂肪酸生物合成等通路。收缩的基因家族的生物学过程只参与了蛋白质磷酸化。
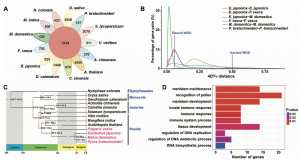
图2 野生枇杷比较基因组分析
3、枇杷种质多样性研究
对26个典型的枇杷栽培种质资源(代表全球不同地区的栽培种质资源)和11个来自中国西南地区的野生种质资源进行了全基因组重测序,平均深度为22.49×,共鉴定出10,978,138个高质量的SNP。系统发育和主成分(PCA)分析表明,野生种质和栽培种质聚为两个不同的组(图3A和B)。野生枇杷的核苷酸多样性(π)(2.28×10−3)高于栽培枇杷(1.44×10−3),LD衰减速度快于栽培枇杷(图3C),期望杂合度和观测杂合度的平均值均显著高于栽培枇杷。进一步利用Structure对种群结构分析显示,当K=2时,所有个体被清楚地细分为野生和栽培种质的两个特定分支(图3D)。

图3 野生和栽培枇杷的群体结构分析
果实品质和色泽是枇杷的重要农艺性状,选择清除分析结果鉴定到283个受选择区域,包含2,381个基因,这些基因功能主要涉及糖、有机酸、脂肪酸、氨基酸、类黄酮、类胡萝卜素和植物激素的生物合成和代谢途径(图4D),参与果实品质与色泽。同时,收到选择的还有果实大小相关的基因如BZR1、BZR2、IAA26、NAC、SAUR32和SAUR72等。另外,这些基因在淀粉和蔗糖代谢、碳代谢、果糖和甘露糖代谢、植物-病原体相互作用、脂肪酸代谢、苯丙素生物合成、类黄酮生物合成、类黄酮生物合成、类胡萝卜素生物合成和植物激素信号转导等通路显著富集。综合这些结果,作者提出了一种独特的枇杷果实品质和果肉颜色的驯化模式。
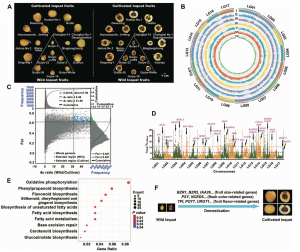
图4 野生枇杷与栽培枇杷选择清除分析
4、枇杷果实中基因表达特性研究
栽培枇杷的果实味道优于野生枇杷,为了确定参与果实品质的关键调控基因,作者对果实发育和成熟的三个阶段(即果实未成熟(GF)、果实转色(CT)和果实成熟(FR))进行了比较转录组分析。在3个发育阶段共表达了23,275个基因,其中5,435个基因表达存在显著差异(图5A和B),这些差异表达基因(DEGs)可能有助于果实的独特特征,主要富集在淀粉和蔗糖代谢、植物激素信号转导、MAPK信号通路等途径。与野生枇杷相比,栽培枇杷在果实发育过程中主要在糖代谢、植物激素信号转导、类黄酮生物合成和类胡萝卜素生物合成等途径富集(图5C和D)。其中,淀粉和蔗糖代谢相关基因均在栽培枇杷的CT和FR期显著上调(图5E)。同时,发现大多数类胡萝卜素生物合成相关基因,在红色果实中表达明显高于白色果实,与长期以来认知一致,即枇杷的红色果肉是类胡萝卜素积累的结果。

图5 枇杷差异表达基因(DEGs)
5、枇杷果实中代谢产物变化研究
为了确定FR阶段的代谢变化,作者基于广靶的LC-MS/MS对野生枇杷和栽培枇杷进行了代谢组研究,共鉴定出1040种代谢物,利用层次聚类分析(HCA)和主成分分析(PCA)对4种枇杷果实的代谢产物进行了分类,发现野生枇杷和栽培枇杷在果实发育过程中代谢产物谱存在显著差异。Jinhua No. 2 FR与GZ-23 FR的代谢组学分析显示,共有371个差异积累代谢物(DAMs),其中149个上调,222个下调(图6D)。但是野生枇杷中一些重要的类黄酮类化合物均显著上调。Huabai No. 1 FR与GZ-White FR比较,共有413个DAMs表现出显著差异,其中上调63个,下调350个(图6C)。其中,鼠李糖、棉子糖、D-葡萄糖、D-半乳糖、D-甘露糖和肌醇在白肉枇杷中显著上调。有机酸中,茉莉酸和2-吡啶甲酸在白叶枇杷中显著上调(图6E)。

图6 野生与栽培枇杷成熟果实的代谢组学分析
最后,为了进一步准确了解成熟果实中转录水平与代谢物变化之间的关系,作者进行了DEGs与DAMs之间的相关分析,其中,变化趋势相同的DEGs和DAMs主要涉及类黄酮、酚酸类、氨基酸及其衍生物、萜类、有机酸、脂类等(图7)。
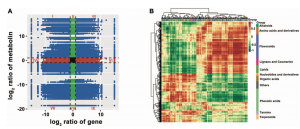
图7 成熟果实转录组与代谢组的相关性分析
总结
野生枇杷为改良品种的驯化和育种研究提供了丰富的遗传资源。作者组装出野生枇杷染色体水平的基因组,比较基因组研究表明枇杷与苹果和梨拥有一个共同的祖先,且在枇杷分化之前就发生了一次WGD事件。基因组重测序结果表明,野生枇杷较栽培枇杷具有较高的遗传多样性,果实质量、大小和叶片颜色相关性状基因在驯化过程中受到了选择。进一步通过转录组和代谢组分析鉴定到野生和栽培枇杷在果实发育不同阶段的DEGs和DAMs,关键差异基因和代谢物主要涉及糖代谢、植物激素信号转导、类黄酮和类胡萝卜素生物合成。这些高质量的参考基因组、重测序、转录组和代谢组分析为阐明枇杷的果实驯化和分子育种研究提供了有利保障。
原文链接:https://doi.org/10.1093/hr/uhac265




 京公网安备 11011302003368号
京公网安备 11011302003368号